Introducción
El sistema
visual nos proporciona información acerca de la localización, forma,
tamaño, textura y movimientos de los objetos. El primer estadio de
procesamiento de esa información tiene lugar en la retina que utiliza luz
emitida o reflejada por los objetos para formar imágenes.
Aproximadamente
la mitad del córtex está dedicada al análisis del mundo visual. El espectro de luz
visible al ojo humano representa sólo una pequeña porción del espectro
electromagnético.
La luz es la variación electromagnética comprendida entre una longitud de
onda entre 400 y 700nm.
·
La longitud de onda (λ). Nos da la percepción del
color, por lo que las diferentes λ se perciben como diferentes colores.
·
Intensidad: percepción de brillo, diferencia intensidades de brillo.
Rango de luz visible: La mezcla
de longitudes de onda en este rango visible a nuestros ojos y a nuestro cerebro
le parece luz blanca, es la luz solar. Cuando impacta sobre los objetos de la
tierra algunas longitudes de onda pueden ser reflejadas por los objetos y otras
son absorbidas por los objetos.
Nosotros sólo percibimos un rango de radiación
electromagnética, denominado espectro o rango de luz visible, que comprende
entre los 400 y 700 NM:
·
Por
debajo de 400 NM nos encontramos con los rayos ultravioletas, los rayos X y los
rayos gamma.
·
Por
encima de los 700 NM nos encontramos con infrarrojos, microondas y ondas de
radio.
Formación de imágenes en
la retina
El ojo posee dos lentes convexas, la córnea y el cristalino que facilitan la
transmisión de energía luminosa y enfocan la imagen sobre la retina. Juntas
desvían (refracción) los rayos de luz para formar imágenes enfocadas sobre la
retina. La curvatura de las lentes y la refracción, hace que la imagen se
invierta en la retina.
Son muy
transparentes para que la imagen sea muy nítida. La retina es sensible a la luz
y transforma la radiación electromagnética en cambios de voltaje, impulsos
eléctricos entendibles por el cerebro, que no pueden interpretar longitudes de
onda.
El ojo contiene
el humor acuoso y vítreo, que son transparentes y permiten formar una imagen
nítida en el fondo de ojo. Si no se conserva esta transparencia hay problemas
visuales.
Organización laminar de la retina
Forma parte del SNC. Posee una organización laminar. Tiene 5 tipos celulares (4
de ellos son neuronas y el tipo celular restante no tiene un origen neuronal,
son los fotorreceptores, que transducen la información, transforman la luz en
impulsos, y no son neurológicos, a diferencia del resto de células de la
retina), y se organizan por láminas. Aunque tengan un origen
neuronal no todas disparan potenciales de acción, solo las neuronas que envían
su axón a las dianas en troncoencéfalo y tálamo.
Hay interneuronas que emiten sus dendritas
lateralmente, establecen conexiones laterales. Hay una vía para el flujo
vertical de información de la retina y hay vías laterales (células horizontales
y amacrinas) para la transferencia de información en la retina.
Interpuestas entre las capas que contienen núcleos
hay dos capas plexiformes que tienen contactos sinápticos: externa e interna.
Las células bipolares, amacrinas,
horizontales, no están mielinizadas y su cuerpo celular es relativamente
transparente. Solo los axones de células ganglionares están mielinizados.
Los elementos sensibles a la luz están
alejados del centro del ojo. A través de la cornea se refractan y atraviesan
todo el humor vítreo y reflejarse en la retina, y además atravesar las capas para
llegar a los fotorreceptores.
Si la luz se distorsiona en el trayecto la nitidez de la imagen se reduce. La
luz debe atravesar todas las capas de células retinianas antes de alcanzar el
segmento externo de los fotorreceptoes. Es importante que todos los medios sean
relativamente transparentes, al igual que las capas celulares.
Los fotorreceptores son muy dependientes de la relación con el epitelio
pigmentario. Los núcleos facilitan la renovación del pigmento que absorbe luz y
tiene un papel importante en el mantenimiento de los discos membranosos de los
fotorreceptores.
Hay 5 tipos celulares, y 3 funcionales:
- Fotorreceptores.
- Interneuronas.
- Células
de proyección. células ganglionares que salen de la retina por el
nervio óptico.
Se disponen en capas
alternas en las que se ubican cuerpos celulares y conexiones sinápticas entre
células.
·
Epitelio pigmentario. La lámina más
externa, más alejada del centro del ojo. No tiene ningún papel en el procesamiento
visual. Tiene melanina que absorbe la luz que cruza la retina que no es
absorbida por los elementos sensibles a la luz. La luz llega a fondo del ojo y
tiene que atravesar todas las capas celulares hasta llegar al elemento
sensible: fotorreceptores. Cuando llega a los fotorreceptores parte de esa luz
podría expresarse retrógradamente y eso degradaría la imagen, para eso hay un
epitelio pigmentario que se encarga de absorber la luz que no es absorbida por
los fotorreceptores. También tiene un papel importante en el mantenimiento, la
nutrición de fotorreceptores, y el reemplazo de subtipos membranosos.
·
Segmento externo de los
fotorreceptores. Tienen
discos membranosos apilados que tienen la molécula que se encarga de absorber,
de fotopigmentos.
- La capa nuclear externa tiene el cuerpo celular de los
fotorreceptores. Tiene el núcleo y todo el material fotorreceptor del que
emanan estructuras que terminan haciendo contacto con las células
horizontales y bipolares los fotorreceptores son células alargadas con: el
Segmento externo y el cuerpo celular
- Capa plexiforme externa. Contiene contacto sináptico
entre las bipolares y amacrinas, horizontales y ganglionares
- La capa nuclear interna tiene cuerpos
celulares de células bipolares, horizontales y amacrinas. Estas
células hacen múltiples conexiones sinápticas laterales, que realizan un
papel regulador.
- Capa plexiforme interna.
- Capa de células ganglionares tiene los cuerpos celulares de
células ganglionariores. Son células de axón largo que
se dirigen a centros de procesamiento en troncoencéfalo y por tanto son
axones de conducción. Únicas que disparan potenciales de acción.
Entre ellas hay capas de conexiones celulares.
En la retina hay conexiones verticales que facilitan el flujo directo y
conexiones horizontales que facilitan interacciones laterales entre los fotorreceptores
y demás células.
Se produce la dilatación de la pupila para observarla con el oftalmoscopio.
Veríamos el fondo del ojo. Es de color pardo, debido al epitelio
pigmentario con melanina, está más externo que el segmento externo de los
fotorreceptores.
Hay una zona pálida, el disco óptico, por el cual salen los axones de las
células ganglionares de la retina, mielinizados, aquí no hay fotorreceptores,
no es sensible a la luz. Además por aquí entran los vasos que se ramifican pro
la superficie interna de la retina.

- Disco
óptico o punto ciego: Región redondeada y pálida, hay muchos
vasos sanguíneos que salen y se ramifican. Además es pálida porque representa
el sitio de salida de los axones de las células ganglionares (que tienen
mielina).En este punto de entrada de vasos
no hay fotorreceptores, esta zona de la retina no es sensible a la luz.
Podríamos pensar que se vería un agujero en el campo visual, sin embargo eso no
es así. El cerebro hace que no veamos ese agujero en el campo visual.
- Mácula:
región amarillenta, estamos viendo la melanina del epitelio
pigmentario, dentro de ella vemos
una zona más oscura, debido a que es más delgada porque los cuerpos
celulares están desplazados hacia los lados, y porque además en esta zona
no hay vasos sanguíneos. Esta zona es la fóvea y tiene más agudeza visual,
por esto enfocamos y movemos los ojos para encuadrar la fóvea sobre el
objeto que miramos.
Dentro
de la fóvea nos encontramos con una parte aún más fina (300 micras), que tiene
aún más agudeza visual, es la foveola.
Hay una zona más oscura, llamada fóvea y en ella la retina es más delgada, y es
la porción que proporciona mayor agudeza visual. Es más delgada porque quita
todas las capas y vasos innecesarios, para que tenga la mayor nitidez. Se usa
como punto de referencia anatómico del centro de la retina. La divide en nasal,
temporal, superior e inferior.
Las capas celulares que están bajo los fotorreceptores están desplazadas
lateralmente eso facilita que la luz llegue a los fotorreceptores con la misma
información óptica.
Cuando se usan ambos ojos no es posible evidenciar que existe este punto ciego.
Cuando utilizamos un sólo ojo el cerebro rellena ese agujero utilizando
información del entorno.
Encontramos dos tipos de fotorreceptores, son diferentes en el tipo de
fotopigmento, en su conectividad y en su distribución:
- Sistema de bastones
- 20
veces más numerosos. De los 126 millones de fotorreceptores de la retina,
120 son bastones.
- Retina
periférica
- Alta
sensibilidad a la luz
- Alta
convergencia.
- Baja
resolución espacial(poca agudeza visual)
- Visión
escotópica (muy sensibles pero se satura más) y acromática.
- Sistema
de conos
- Retina
central (fóvea)
- Baja
sensibilidad a la luz, necesita más fotones de luz que los bastones
- Reducida
convergencia
- Alta
resolución espacial (alta agudeza visual)
- Visión
fotópica y cromática (en color, proporcionan información específica sobre
las longitudes de onda)
En el segmento externo hay discos membranosos afilados, donde hay fotopigmentos
importantes para la transmisión.
Bastones
Los bastones son tan sensibles a la luz por la cantidad de fotopigmentos,
tienen un segmento externo (cilíndrico) más alargado que los conos, donde
hay muchos discos membranosos con fotopigmentos que los hace más
sensible. Para que la luz sea detectada por los conos se necesitan unos 100
fotones.
Son muy convergentes, se va a recibir mucha información de muchos bastones.
Aunque disminuye la discriminación aumenta la sensibilidad, menor intensidad
para generar una respuesta.
Funcionan bien en condiciones de iluminación tenue porque con demasiada luz,
los bastones se saturan y dejan de responder por lo que los bastones son
importantes en condiciones de luz tenue como puede ser el crepúsculo o la
noche.
Conos
Tienen un segmento externo cónico y tienen menos discos membranosos. Los conos
por el contrarios como son menos sensibles y menos convergentes, les capacita
para ser un sistema con mayor resolución y, además, proporcionan una luz en
color y una visión ecológica, porque en su membrana se expresan, 3 tipos de
fotopigmentos, sensibles a una porción distinta del espectro lumínico. Cada
cono tiene un fotopigmento diferente (en los bastones solo hay un tipo de
fotopigmento). En la retina hay un cono por célula bipolar por célula
ganglionar por lo que hay 0 convergencia.
En los discos membranosos de los fotorreceptores hay fotopigmentos que son
moléculas sensibles a los fotones. Está formado por dos porciones:
- Una proteica (opsina,
atraviesa 7 veces la membrana) es la que varía entre los diferentes conos
(conopsina) y bastones (rodopsina) tiene una molécula más pequeña
derivado de la vitamina A que es el cromóforo, que es el que absorbe la
luz. Está unido a una parte proteica (transduce la señal lumínica) y por
la parte que absorbe luz que es el retinal.
El retinal
tiene varias conformaciones isoméricas, dos importantes en la traducción.
El cis retinal es cuando no ha absorbido luz, cuando lo hace, se transforma
en trans retinal, haciendo que la opsina se active, activando elementos de
traducción.
Fototransducción
Cuando la molécula de fotopigmento absorbe color, la rodopsina (en los
bastones), en el cono es la conopsina y hay 3 tipos diferentes. Este es el
único sistema en el que la luz hiperpolariza al fotorreceptor.
En la oscuridad los fotorreceptores están despolarizados y liberan de forma
tónica glutamato, porque están parcial despolarizados porque hay canales de Na
abiertos, estos canales están regulados por nucleótidos, de GMPc, que tiene
niveles elevados en oscuridad, por lo que hay flujo continuo de Na hacia el
interior.
Cuando se proyecta luz, se activa el fotopigmento y el retinal que es el que
absorbe el protón, se disocia de la opsina. La opsina está activa y activa a
proteínas G (transducciona) esta proteína G releva la señal a una enzima
efectora, la fosfodiesterasa que rompe el gen específico que hay en el
citoplasma de los fotorreceptores, cayendo los niveles de GMPc.
Si los niveles del gen cae, los canales dependientes del gen específico se
cierran y la membrana se hiperpolariza. Este mecanismo es igual en conos y
bastones. En los conos depende la longitud de onda de la luz para hiperpolarizarse
o no, en los bastones no.
En todos los
sistemas sensoriales que hemos visto y veremos, la estimulación, la
transducción del estímulo, casi siempre conlleva despolarización de la
célula. El caso de los fotorreceptores es una excepción, cuando transducen el
estímulo, lo que hacen es interpolar, es decir, se comportan de forma
contraria, causa hiperpolarización.
A continuación
vamos a explicar el proceso que se lleva a cabo en los bastones, siendo
aplicable también para los conos.
En condiciones de
oscuridad el retinal se encuentra pegado a la rodopsina y la célula está
parcialmente despolarizada, su potencial de membrana sería unos -40 mV y
estaría liberando continuamente glutamato. Está despolariza porque hay
canales de sodio abiertos, estos canales de sodio están regulados por nucleótidos.
En condiciones de oscuridad los niveles de GMPc intracelular están elevados.
Cuando se
ilumina: el fotopigmento absorbe fotones, la rodopsina se disocia, el retinal
cambia de cis a trans y se disocia de la rodopsina, eso hace que la parte
proteica quede activada. Una vez activada va a activar a una proteína G
(actuará por tanto como receptor metabotrópico) que activa a una enzima
(fosfodiesterasa) que baja la cantidad de GMPc. Si caen los niveles de GMPc
acaban cerrándose los canales de Na+ y por tanto se produce una
hiperpolarización. La consecuencia es que en presencia de luz la membrana del
fotorreceptor se hiperpolariza.
|
Sensibilidad espectral
de conos y bastones
Los tres tipos se diferencian por el tipo de opsina que hay en el fotopigmento que
es el que afina el espectro de absorción y hace que absorba determinadas
longitudes de onda y no otras.
El fotopigmento es el retinal (cromóforo) junto con la opsina (parte proteica;
distintas isoformas). En la retina hay 4 isoformas de la opsina que
confieren distinta sensibilidad espectral a la molécula de fotopigmento (4
tipos de fotopigmento).
El retinal absorbe la luz, pero es la opsina la que hace que el retinal
absorba una determinada longitud de onda.
El fotopigmento de los bastones es la rodopsina de la cual hay un solo tipo, lo
que da una visión monocromática.
El fotopigmento en los conos es al conopsina, de la cual hay 3 tipos lo que nos
proporciona una visión tricromática.
·
Conos para el azul (conopsina absorbe λ corta) 420NM.
·
Conos para el verde (conopsina absorbe λ intermedia) 530N.
·
Conos para el rojo (conopsina absorbe λ larga) 560NM.
Si se activan los bastones, la percepción visual que tiene es
monocromática. Cuando se activan los conos, nuestro cerebro percibe luz en
color. D e hecho, el color de la luz depende del nivel relativo de actividad
del conjunto de estos tres tipos de conos.
Hay tres tipos de conos que absorben las 3 longitudes de onda, el cerebro para
asignar un color a la percepción hace una lectura de el nivel relativo de
actividad de estos tres conjuntos de conos de forma que si los tres tipos de
conos están activos simultáneamente la luz percibida sería blanca., si están
activos únicamente los conos azules y no los verdes y los rojos, la luz
percibida será azul. Si se activan los verdes y los rojos y no los azules, la
luz sería amarilla y así sucesivamente.
Los colores son percepciones creadas por el cerebro en base a la longitud de
onda que llega a la retina.
Si una de los tipos de las opsinas, expresadas en los conos, falta. Se produce
el daltonismo (visión dicromática), la cual es muy frecuente y
provoca escasas consecuencias. Los genes para unos de estos tipos de opsina o
no existe o haya un híbrido modificado.
Hay una capacidad alterada para discriminar colores, sobre todo verde y rojo. Estos
dos genes son homólogos en la secuencia y están muy próximos entre sí en el
cromosoma X, lo cual explica que el daltonismo sea más frecuente en hombres que
en mujeres. Puede ser que en la meiosis se produzca una alteración en estos
genes.
El cerebro obtiene
información acerca del color de los objetos comparando los niveles relativos de
actividad de los 3 tipos de conos.
Distribución diferencial de conos y bastones en la retina
En la mayor parte de la porción periférica de la retina dominan los bastones,
los conos representan una minoría (6 millones de los 126) y están concentrados
fundamentalmente en la región de la fóvea, región de unos 2 mm. En esa región
de la fóvea no hay bastones prácticamente. En la foveola (punto central de la
fóvea) no hay ningún bastón.
El segmento externo de los conos en la fóvea es reducido, de manera que se
pueden apilar siendo su densidad muy alta.
En la fóvea el segmento externo de los fotorreceptores tiene un diámetro
muy reducido, eso hace que estén muy empaquetados. A medida que nos vamos a
posiciones más excéntricas, el segmento externo aumenta de diámetro y están
menos empaquetados. Es
como si tuviéramos dos retinas, una que sirve para ver en color y alta resolución
y poco sensible a la luz (porción central), y otra retina muy sensible a la luz
pero que tiene muy poca capacidad de resolución especial (bastones).
Convergencia en los sistemas de conos y bastones
Las células que vienen de la fóvea reciben información de una sola célula
ganglionar.
El sistema de conos es menos convergente (cada cono
manda la información a una sola neurona bipolar), lo que hace que tengan menos
sensibilidad pero una alta capacidad de resolución espacial.
En los bastones hay una convergencia mayor, por lo que sus sensibilidad a la
luz es mayor, teniendo menor resolución espacial.
A medida que disminuye la convergencia aumenta la
resolución pero baja la sensibilidad (con la convergencia se suman pequeñas
señales que conforman una mayor sensibilidad). Aumenta la resolución espacial
pero no es lo mejor si se quiere sensibilidad a la luz, si esto es lo que se
quiere ha de ser convergente.
Al fijar la vista en un punto que brilla se puede tener la percepción de otro
punto, más tenue, lo cual es la percepción de la porción periférica de la
retina. El centro de la retina es donde tiene más resolución (pixeles en una
cámara fotográfica) y la periferia, donde menos.
La agudeza visual se reduce en un 75% a sólo 6% de la línea central de
visión.
Resumen de las propiedades diferenciales
de conos y bastones
|
Bastones
|
Conos
|
Distribución
|
Periférica
|
Fóvea
|
Número
|
Mayor (20x)
|
Menor
|
Cantidad de fotopigmento
|
Mayor
|
Menor
|
Convergencia
|
Aproximadamente 120/fibra n. óptico
|
Aproximadamente 8/fibra n. óptico
|
Sensibilidad
|
Mayor
|
Menor
|
Discriminación espacial
|
Menor
|
Mayor
|
Responde mejor a
|
Luz difuminada
|
Luz directa
|
Visión
|
Escotópica acromática
|
Fotópica cromática
|
Interacciones sinápticas laterales
Estas interacciones proporcionan inhibición lateral (GABA, glicina) de los
circuitos, a través de células horizontales y células amacrinas.
Hay
dos vías para la transferencia de información desde un fotorreceptor a una
célula bipolar dependiendo del tipo de información que sea.
Hay una vía que hace contacto por células bipolares
y hay vías indirectas mediadas por estas conexiones laterales en las que
intervienen las células horizontales. La actividad de la célula bipolar puede
estar regulada por fotorreceptores de los que recibe contacto directo y también
por fotorreceptores adyacentes. Es decir, hay dos vías posibles:
Los fotorreceptores liberan glutamato que es el NT
excitador del SNC. En el caso de células bipolares, hay algunas que son
excitadas por glutamato y otras inhibidas por glutamato. Es decir que el
glutamato que liberan los fotorreceptores puede activar a unos tipos de células
bipolares e inhibir a otras. La diferencia es el receptor para glutamato que
expresan las células bipolares. Las conexiones en células ganglionares son
siempre activadoras (también con glutamato).
Por el contrario entre células amacrinas y
horizontales utilizan GABA y glicina que son dos NT inhibidores.
Transmisión de la
información de las células de la retina
La mayor parte de las células de la retina no crean potenciales de acción, las
únicas que lo hacen son las células ganglionares, las de proyección, las que
envían la información al encéfalo, el resto, la mayor parte, no.
La información la trasmiten, al hiperpolarizarse, se transfiere en forma de un
potencial graduado, local, a las células ganglionares. El potencial del
receptor se propaga por toda la membrana, y llega hasta las vesículas de
glutamato que se liberan. Por lo que no necesita potenciales de acción ya que
son células pequeñas, con este potencial de receptor, tienen suficiente. Es un
potencial graduado, que se atenúa, y al ser pequeñas no tiene problema.
Campos receptores
Las células bipolares y ganglionares poseen campos receptivos circulares con
antagonismo centro-periferia.
El campo receptor de una célula del sistema visual, una célula de la retina por
ejemplo. El campo receptor en una célula retiniana es el área de la retina que
monitoriza, dónde hay fotorreceptores que actúan directa o indirectamente sobre
la célula.
El campo receptor de las células de la retina tiene dos características:
- Suelen
ser circulares
- Muestran
dos regiones diferenciales, una región central circular y una porción
periférica a modo de anillo alrededor de ese centro.
Los fotorreceptores del campo receptivo contactan directamente con la
célula ganglionar. A través de conexiones horizontales de las células bipolares
se producen conexiones indirectas.
De una región adyacente de la retina se puede recibir información también. Por
lo que hay dos regiones, centro y periferia, son circulares. La respuesta que
induce la estimulación por luz del centro del campo receptivo se cancela cuando
simultáneamente se estimula la periferia por lo que hay antagonismo.
Si yo ilumino el centro del campo selectivo, las células se despolarizan. Es
una respuesta graduada. Si en vez de iluminar el centro, ilumino la periferia
la célula ganglionar se hiperpolariza. Esta respuesta antagónica esta mediada
por las conexiones laterales, que ocasiona las células horizontales. Esto
ocurre en la lámina interna, en la externa están las amatrinas que también
provocan inhibición lateral.
La respuesta a la luz en un fotorreceptor es hiperpolarizante, liberan
glutamato, el cual es un neurotransmisor excitador, casi siempre. Si se cumple
esto, al iluminar el segmento externo del foto la membrana se hiperpolariza y
se reduce la liberación de glutamato desde sus terminales. Esto en el potencial
de membrana de la células diana (bipolar), provoca que se hiperpolarice, es un
cambio en el PM que va en la misma dirección. Pero en realidad, se despolariza
al liberarse menos glutamato, porque aunque es NT excitador, en las sinapsis lo
que hace es una acción inhibidora. Son canales de tipo alfa, que son catiónicos,
y se despolariza.
Otras veces es metabotrópico, y se abren o cierran canales lo que hace que
pueda ser hiperpolarizante.
Los fotorreceptores de la retina hacen contacto con dos tipos de células
bipolares: en unas es despolarizante y en otras hiperpolarizantes. Se
diferencia en el tipo de receptor que expresan. Todos los fotorreceptores
envían información de manera simultánea a ambos tipos de células bipolares.
Tipos de células ganglionares según su respuesta a la iluminación del
centro del campo receptivo
Las células ganglionares tiene campos receptivos circulares que muestran un
centro y una periferia, habiendo un antagonismo campo-periferia.
Una célula ganglionar es el único tipo celular de la retina que genera
potenciales de acción entonces podemos ver cómo responde registrando en el
nervio óptico los axones.
Tienen campos receptivos circulares que muestran un centro y una periferia y,
además, hay antagonismo centro periferia.
En basa a la respuesta de estas células cuando se ilumina el centro del campo
receptivo:
Ocurre también en las células bipolares
pero con respuesta graduada no con potenciales de acción.
La actividad de
fotorreceptores adyacentes también puede modificar la actividad de estas
células, en este caso lo que hace es antagonizar la respuesta.
Esto lo que hace
es tornar a estas células como células muy sensibles al contraste de
luz-oscuridad, es decir, van a responder muy bien si un borde luz-oscuridad
cae en su campo receptivo. (Pero los hace malos para transmitir al cerebro
información sobre niveles de luz absolutos).
Si tengo un objeto con forma cúbica en la mesa, la luz que viene de
arriba hace que cada una de las paredes del objeto refleje luz en una
dirección, los bordes de esas paredes son bordes donde hay contacto
luz-oscuridad, eso es lo que está enviando información acerca de la forma de
ese objeto. Es el patrón de contraste de luz en los bordes es lo que da la
forma de los objetos, el que dice qué objeto es.
La
respuesta es débil cuando el patrón de iluminación de sus campos receptivos
(CRs) es uniforme y es mayor si hay diferencias de iluminación (contraste de
luz; bordes luz-oscuridad) en el interior de sus CRs.
Cada
célula ganglionar envía señales a ambos tipos de neuronas bipolares (tanto a
las que se despolarizan con la luz, como a las que se hiperpolarizan).
Cada
fotorreceptor envía información a los dos tipos de células bipolares, son
como dos canales paralelos. Las células bipolares que se despolarizan con luz
hacen contraste con células ganglionares, esto hace que las células
ganglionares se despolaricen con luz, es decir va a haber células bipolares
que se despolaricen y que se hiperpolaricen con luz. [Cada fotorreceptor
envía señales a los dos tipos de células (2 canales paralelos, las
ganglionares de centro ON y las de centro OFF)].
|
La respuesta es débil cuando el patrón de iluminación de sus CRs es
uniforme y es mayor si hay diferencias de iluminación (contraste de luz, bordes
luz-oscuridad) en el interior de sus CRs. Cada fotorreceptor envía señales a
los dos tipos de células (dos canales paralelas).
Cuando aplico luz a las dos regiones del campo receptivo hay una
cancelación centro-periferia y en este caso no tengo cambio en la
actividad de disparo de las células.
Estas células son muy buenas para responder a la luz cuando los patrones
luz-oscuridad caen sobre el campo receptivo pero no son buenas para informar
sobre los niveles globales de iluminación.
Son muy buenas para detectar contrastes, borde de luz. Esta configuración del
campo las hace buenas para estas diferencias de luz.
Al igual que con las células bipolares, las células ganglionares las hay
de dos tipos, ya dichos, y todas las regiones de la retina están monitorizadas
por ambos.
- Si se
mueve un borde de oscuridad a través del campo receptivo de las células:
todo el campo iluminado.
- Si la
sombra cubre parte de la periferia, si es ON, no se contrarresta
totalmente, y la tasa de disparo aumenta.
- Si se
cubre el centro, al ser ON, la células de silencia.
- Si se
cubre totalmente, la rama de disparo apenas se diferencia de la anterior.

Esto no es bueno para informar acerca de
diferencias de la cantidad de fotones que llegan a la retina, sino par a la
presencia de bordes luz oscuridad, esto es lo que se exagera, las sombras.
A más intensidad, mayor cantidad de luz. Lo relevante son los patrones de
contraste, patrones de diferencia de brillo, que es lo que nos permite
reconocer, analizar el contorno de los objetos.
Si se proyecta un borde luz-oscuridad en la retina (es un borde estacionario).
No lee la actividad de las células ensombrecida o totalmente iluminan sino
leyendo los cambios que se dan en las células que están en el borde, puesto que
son las que se modifican, ya que si están en sombra o luz total no cambian. Se
modifican porque tienen diferenciado, en función de luz u oscuridad, su centro
y su periferia.
La percepción de la luz es relativa
Nuestra retina no está especializada en detectar cambios en los niveles
absolutos de iluminación, sino variaciones espaciales de tipo local. Niveles
absolutos de iluminación, sino variaciones espaciales de tipo local.
La señal que sale de la retina exagera las diferencias, las amplifica. Por eso
da la sensación de que cada cuadrado es de una tonalidad, y esto es generado en
el cerebro, por la diferencia entre los fondos. La retina destaca contrastes.
Clasificación de células ganglionares según conectividad, apariencia y
propiedades fisiológicas
|
Células M
|
Células P
|
Porcentaje relativo
|
 10 10
|
80
|
Localización en retina
|
|
Mayor densidad en fóvea
|
Velocidad de conducción
|
15 m/s
|
6 m/s
|
Resolución temporal
|
Muy alta (>60Hz)
|
Alta (>30Hz)
|
Información cromática
|
Prácticamente ninguna, reciben pocas
entradas de conos.
|
Si (responden a λ específicas)
|
Información de bastones
|
Si
|
A veces
|
Resolución espacial
|
Menor (>campo receptor)
|
Mayor (<campo receptor)
|
Zonas de proyección
|
NGL magnocelular
|
NGL parvocelular
|
En la retina hay células ganglionares de
tipo M y de tipo P, las cuales son las más abundantes y hay mayor densidad en
la fóvea. La resolución temporal (Cuando algo cambia en la escena visual) es
que son capaces de responder, cuando un estimulo luminoso se apaga y enciende
muy rápido. Las M son más grandes.
La información cromática es la del color.
Las células P reciben información de los conos. La resolución espacial donde es
mayor debido a que el campo receptor es menor. La célula más directamente
implicada en la percepción de formas detalladas, detalles finos son las células
P, puesto que está relacionado con la resolución espacial. Y en la detección
del movimiento son las M, puesto que tiene que ver con la resolución temporal.
Y el color, las mejores son las P, debido
a que reciben información de los conos.
Terminan en distintas regiones del tálamo. Están segregadas. En el tálamo y en
la corteza visual.
La vía M es importante para la percepción de movimiento y la P para la
resolución de detalles pequeños y color.
En cada tipo hay células de centro ON y de centro OFF.
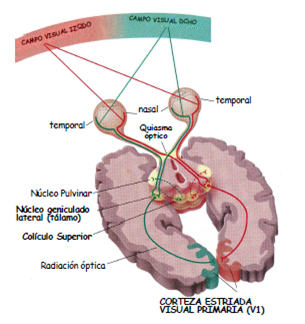
Organización general de las vías visuales
El
campo visual lo podemos definir como el espacio que vemos cuando estamos con la
mirada fija al frente. La zona del campo que se ve al tener los dos ojos abiertos. Se le traza
una línea vertical y se divide una parte izquierda y una derecha.
La fóvea divide la
retina respecto a dos ejes, en 4 partes:
o
Hemirretina nasal (más medial).
o
Hemirretina temporal (más cerca del hueso temporal).
El campo visual izquierdo lo ve la porción
temporal de la retina derecha y la porción nasal del izquierdo, mientras que el
CV derecho, es detectado por la porción temporal de la retina izquierda y la
porción nasal de la derecha. El campo visual derecho lo vemos en la hemirretina
nasal del ojo derecho y con la hemirretina temporal del ojo izquierdo.
Los axones se agrupan
para formar en N. óptico, que tiene una zona llamada quiasma óptico, en el que
hay un cruce de algunos axones ganglionares, de aquellos que provienen de la
porción nasal de cada retina. Los axones de las células ganglionares en la
porción nasal de cada retina cruzan al lado contralateral en el quiasma óptico.
A partir del quiasma hay una representación compleja del campo visual del campo
contralateral. Cada hemisferio cerebral procesa información visual del hemicampo
visual contralateral.
En el tracto óptico izquierdo está el CV derecho y en el tracto derecho
está el CV izquierdo.
Las mitades del tálamo reciben información de lado contralateral.
Dianas subcorticales de los axones de las células ganglionares
La más importante de las proyecciones ganglionares es el núcleo geniculado
externo, del tálamo (99%). El núcleo talámico, donde se procesa la información
que se envía al córtex para percepción visual es el núcleo geniculado lateral,
de donde se manda a la corteza estriada del lóbulo occipital.
Hay otras zonas, como el hipotálamo, cuya información es importante para
controlar los ritmos circadianos.
Hay proyecciones al área precentral del mesencéfalo, que controlan de manera
refleja la pupila y el cristalino, tamaño y forma respectivamente. Para regular
la cantidad de luz, y ver objetos cercanos y lejanos.
Y proyecciones al colículo superior, información para coordinar movimientos
circulares rápidos de los ojos y la cabeza.
Además de estas dianas subcorticales, hay otras
dianas subcorticales a las que llegan axones ganglionares. (Aunque el núcleo
geniculado lateral es el más importante):
- Al
hipotálamo para regular los ritmos
circadianos que oscilan siguiendo las fases de luz-oscuridad.
- Al
Área pretectal del mesencéfalo
(cerca del colículo superior), para controlar los reflejos pupilares. Cambia el
diámetro de la pupila en función de la cantidad de luz que incide en los ojos
(mediante dos músculos).
- Al
colículo superior, donde hay un mapa
del campo visual contralateral, que controla los movimientos oculares rápidos.
Reflejo fotomotor
(pupilar)
En condición de luz no muy elevada, se ilumina una de las retina, la pupila se
contrae en ese ojo y en el del otro ojo, es la respuesta directa y la
consensuada.
Hay axones que se proyectan al área pretectal del mesencéfalo, donde hay
neuronas que proyectan bilateralmente al núcleo accesorio promotor, que tiene neuronas
preganglionares parasimpáticas, cuyos axones discurren el N IIIà después a unas posganglionares
parasimpáticas, y alcanzan el músculo esfínter de la pupila. Y cuando se
ilumina de las retinas se desencadena una respuesta de cierre pupilar en el otro
ojo. Se suele afectar en traumas.
Organización del núcleo geniculado lateral
Las entradas ganglionares M y P se mantienen segregadas en este núcleo y en
áreas corticales, terminan en diferentes capas.
En el núcleo geniculado lateral tenemos una serie de capas que se nombran del 1
al 6.
- Las
capas 1 y 2 se llaman magnocelular y reciben entradas de las células
ganglionares tipo M.
- Las 4
más dorsales (3, 4, 5 y 6) reciben entradas de células ganglionares tipo
P. La segregación funcional de ambas vías se preserva o parvocelulares.
Y, además, cada capa recibe entrada de un único ojo (2, 3 y 5 vs 4 y 6), 2,
3 y 5 recibe entradas de la hemirretina temporal homolateral y las 1, 4 y 6 de
la hemirretina nasal del ojo contralateral, por lo que se mantienen segregadas
las entradas y los atributos de la información visual a este nivel. Sabemos que
por cada tracto óptico van fibras nasales contralaterales y fibras temporales
ipsilaterales que terminan en distintas capas. Las fibras temporales
ipsilaterales terminan en las capas 2, 3 y 5 mientras que las fibras nasales
contralaterales terminan en las capas 1, 4 y 6.
Distintos atributos de un estímulo se procesan de manea separada.
Además, otra característica del patrón de aferencia es que las proyecciones son
ordenadas, de forma que hay un mapa retinotópico, completo del campo visual
contralateral, en cada lado del núcleo geniculado, el derecho y el izquierdo.
No todas la regiones de la retina están mapeadas en la misma escala, la mayor
parte esta ocupara por regiones de la fóvea y zonas cercanas a ella, es casi
más de la mitad. Están hiperrepresentadas. Proyecciones ordenadas (en cada NGL
existe un mapa retinotópico completo de la mitad contralateral del campo
visual).
Los campos receptivos son concéntricos y muestran antagonismo centro periferia
(similares a los de las células ganglionares de la retina).
Corteza visual estriada y extra-estriada
La información que llega al tálamo está fragmenta y envía la información a la
corteza.
·
V1: corteza visual primaria à corteza estriada (Área 17 de Brodmann).
·
V2 y V3: corteza visual secundaria. Forman un cinturón que rodea a V1.
·
V4: color y forma. Ocupa una posición ventral.
·
V5 (MT): movimiento. Ocupa una posición más dorsal.
Desde V2 a V5: corteza extra-estriada.
Los axones talámicos envían informa a la corteza visual primaria, la
información se procesa en V1 y luego, se continúa procesando en (corteza
visual secundaria) V2, V3 y en áreas corticales de asociación, V5 (movimiento)
y V4 (color y forma).
A la corteza visual primaria del área 17 también se le llama corteza estriada
porque es muy amplia y en preparaciones histológicas se ve como banda clara muy
amplia y al resto de áreas se le denomina corteza extraestriada.
En
corteza visual primaria y en corteza visual de orden superior, esa segregación
se mantiene (ocurre una extensión de lo que ocurría en el NGL), esas dos vías
se siguen manteniendo separadas, en algún momento cabe esperar que la
información se integre. (Vías de las P y M).
La corteza estriada
(corte visual primario) tiene este nombre debido a que en ella hay una estría
de sustancia blanca, que es debida a los axones de las fibras aferentes.
Corteza visual primaria (V1). Representación del campo visual

Retinotopía: Proyecciones ordenadas
aferentes a V1 crean en cada hemisferio un mapa retinoóptico completo de la
mitad contralateral del campo visual.
Mayor superficie dedicada a la fóvea y áreas circundantes (aproximadamente la
mitad de V1).
Representación de V1: La mayor parte está en la superficie medial, hay un mapa
completo de la retina y del CV, en la superficie de V1. EL cerebro derecho,
procesa información del lado izquierdo y a la inversa.
Por lo que se procesa información de manera contralateral. El hemisferio del
lado derecho procesa la información del lado izquierdo.
Además, por encima de la fisura calcarina se procesa información del hemicampo
visual inferior, y en la zona inferior, del hemicampo superior.
No todas las regiones de la retina están representadas en la misma proporción,
hay más de la fóvea, está hiperpresentada en la corteza visual primaria. Esto es lógico
debido a que hay una mayor densidad de receptores y una menor convergencia
(esto es parecido a lo que ocurría con los dedos en la corteza
somatosensitiva).
Campos receptivos de neuronas de V1
El campo receptivo de las células V1 es algo más complejo y grande,
además encontramos que a estos niveles las neuronas responden a bordes
iluminados, a segmentos, a estímulos con bordes lineales.
La mayor parte de las células V1, en casi todas las capas hay células que
responden a estímulos que poseen una orientación específica y el estímulo que
mejor las activa es un borde orientado de luz. Esto nos permite analizar el
contorno de los objetos y empezar a reconstruir su forma.
Tipos de neuronas en V1 según el tipo de estímulo al que mejor responden
Las células de la capa 4c son de gran importancia porque reciben entradas
desde NGL (neuronas ganglionares...). Es la capa más gruesa.
Las de esta capa 4 envían información a neuronas estrelladas.
Por encima y por debajo de la lámina 4c hay células de dos tipos
- Simples. Responden a barras o segmentos lineales con una
determinada posición y orientación. Cuando tiene otra orientación o se
sale de esa zona central ya no responden.
- Las complejas
responden a estímulos orientados, pero la posición del estimulo en el
campo no es tan determinante e incluso responden mejor cuando el estimulo
se mueve.
Los campos receptivos son bastante distintos a los
que veíamos en retina y en núcleo geniculado lateral, estos eran campos
circulares. Aquí vemos que la mayor parte de las células de V1 ya no responden
a puntos de luz. Ahora responden a barras de luz, segmentos lineales o bordes
con una orientación determinada. Tienen campos receptivos elongados y más
concretos. Tiene también de campo ON y OFF.
Distintos
campos, zonas de la corteza responden a barras de luz con orientaciones
específicas y localizadas en un área determinada en el campo visual. Se cambia
la orientación de la barra de luz. La respuesta de la célula horizontal es más
intensa.
A estas columnas o
barras, se las ha llamado columnas de orientación.
Hay preferencias de
orientación según la forma, de forma que a medida que avanzamos en la corteza
vemos zonas más sensibles a distintas orientaciones y si sumamos todos se
formaría la vuelta completa. Existen células simples que tienen respuesta
específica a barras con una posición y orientación y células complejas con respuesta
a barras con una determina orientación y/o movimientos en sentido específico.
La capa 4C es donde llegan esos axones de la retina.
Las células por encima y debajo de capa 4C responden a estímulos orientados.
Organización modular de
V1
Entradas talámicas a capa 4 segregación de entradas 8 (se procesa por separado
toda la información que viene de un punto concreto de la retina):
El córtex visual primario tiene una organización columnar. Tiene 3 formas
de organización:
- Columnas de orientación. Todas las células de la
columna responde a estímulos que tienen la misma orientación vertical.
Tienen campos receptivos solapados. Excepto por 4c que los campos
receptivos son circulares.
- Columnas de dominancia ocular. Son columnas de células cuya
característica es que recoge información de un solo ojo, de manera
alternante.
- Blobs. Limitados a las capas 2 y 3.
Las células no tiene preferencia de orientación solo responden al color,
su campo receptivo es circular.
Esto es lo que hace es procesar por separado distintos aspectos de la
imagen.
Hipercolumnas en el campo visual córtex visual primario es una espacio de corteza,
pequeño (1mm cuadrado), que tiene una secuencia completa de columnas de
orientación, tiene columnas de dominancia ocular y varios blobs. Analiza toda
la información que vine de una pequeña aparte del campo visual, de la retina.
La información se procesa de forma fragmentada.
Sería un conjunto de hipercolumnas. Las primeras células que reciben
información binocular en V1.
Vías paralelas de procesamiento en la corteza extra-estriada
Desde V1, V2 y V3 distintos atributos de la escena visual son procesados en
paralelo en dos vías separadas de M y de P.
En el córtex se ven dos vías:
- Vía dorsal (procesamiento de la información que va
por V2 y V5). Sale de V5 (MT). Está muy cerca del córtex parietal
posterior. En esta vía se procesa información relacionada con la visión
especial, movimiento, profundidad, distancia relativa, localización, como
lo vemos. Es suministrada por la vía M. Las vías extraestriadas forman
parte de esta vía.
- Vía ventral. Discurre por V4 (zona cortical especial), y
confluye en el córtex temporal izquierdo. Se procesa el color, y la
discriminación detallada de formas que vemos.
Son distintos atributos
que se mantienen segregados a nivel cortical. Al final esto se unifica.
Esto se sabe por las consecuencias de las lesiones. Una lesión en un punto de
la vía dorsal, puede desencadenar un déficit en la percepción del movimiento en
el CV contralateral, agnosia al
movimiento (lesión en V5). Se puede desencadenar una agnosia a la
profundidad (lesión en V5).
En la vía ventral puede provocar que se deje de percibir colores, pero no es
daltonismo. A ese déficit se le denomina acromatopsia (lesión en V4).
Puede haber déficit por reconocimiento de objetos, agnosia a objetos o
en el reconocimiento de caras (prosopagnosia, lesión en corteza temporal).